1. Vorbemerkungen
Über den Artkomplex Osmoderma gibt es in Europa und speziell in Mitteleuropa seit einigen Jahren einen regen wissenschaftlichen Diskurs (Tauzin 1994a, 1994b; Krell 1996; Gusakov, 2002, Ranius et al. 2005, Audisio et al. 2007 u. a.). Nach Auffassung von Tauzin (1994 b) existiert im westlichen und südwestlichen Mitteleuropa Osmoderma eremitum und im östlichen Mitteleuropa Osmoderma lassallei septentrionale. Krell (1996) vertritt die Ansicht, dass es sich lediglich um geographische Rassen handelt, da er Übergänge im Bau der Parameren der männlichen Genitalien festgestellt hat und von einem Überlappungsgebiet in Sachsen und Bayern ausgeht. Das Resultat einer mokelularen Analyse von Audisio et al. (2007) ist u. a. eine Unterscheidung in zwei „Arten“: im westlichen Mitteleuropa O. eremita Scopoli, 1763 und im östlichen Mitteleuropa O. barnabita Motschulsky, 1845, wobei es in Deutschland eine Überlappungszone geben soll. Gusakov (2002) bezeichnet diese osteuropäische „Art“ als O. coriarius De Geer, 1774.
Die Diskussion ist naturschutzpolitisch und -rechtlich relevant, da mit der Flora-Fauna-Habitat-Richtlinie der EU (1992) je nach Auffassung nur eine Art (O. eremita) und nicht O. lassallei bzw. O. barnabita bzw. eine Art mit zwei Unterarten (O. eremita eremita und O. eremita lassallei bzw. Osmoderma eremita barnabita) den hohen europaweiten Schutzstatus als „prioritäre Art“ des Anhangs II der FFH-Richtlinie erhält.
Im Zuge der nacheiszeitlichen Vegetationsentwicklung und Ausbreitung der Laubmischwälder nach Norden (Hewitt 1999) haben sich vermutlich auch Osmoderma spec., die während der Eiszeit in südeuropäischen Refugien überdauert hatten, gen Norden ausgebreitet und Mitteleuropa wiederbesiedelt. Wie diese Wiederbesiedlung ablief, lässt sich nur mutmaßen: wahrscheinlich um die Alpen herum, einerseits aus Südwest- und andererseits aus Südosteuropa. Unklar ist, ob und wann es zur Überlappung der Areale in Mitteleuropa gekommen ist und ob es zu „natürlichen Kreuzungen“ kommt.
Audisio et al. (2007) empfehlen eine Kombination aus diagnostisch morphologischen Merkmalen und Analyse bestimmter Nukleotidsequenzen. In jüngster Zeit wird versucht, eine Klärung durch DNA-Analysen zu erreichen (T. Martschei, mündl. Mitteilung).
Die morphologischen Arttrennungsmerkmale v. a. anhand von Profilzeichnungen der Genitalstrukturen des Männchens, die Tauzin (1994a 1994b) und Krell (1996) veröffentlicht haben, werden bei Rössner (2012) (Abb. 08-1) und bei Schaffrath (2003a) gegenübergestellt (Abb. 08-2).
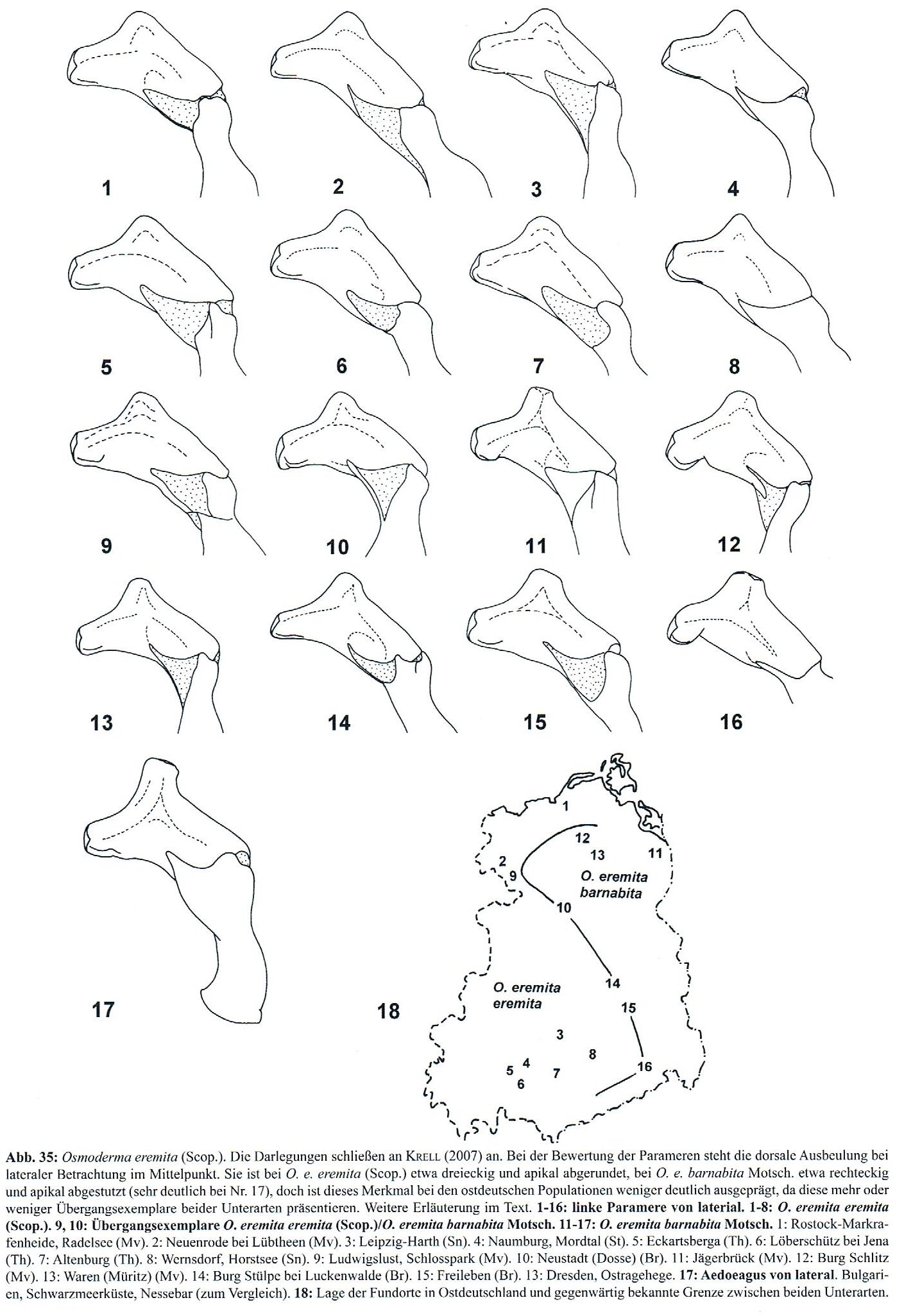 Abb. 08-1: Übersicht zum Bau des männlichen Genitals aus: Rössner (2012)
Abb. 08-1: Übersicht zum Bau des männlichen Genitals aus: Rössner (2012)
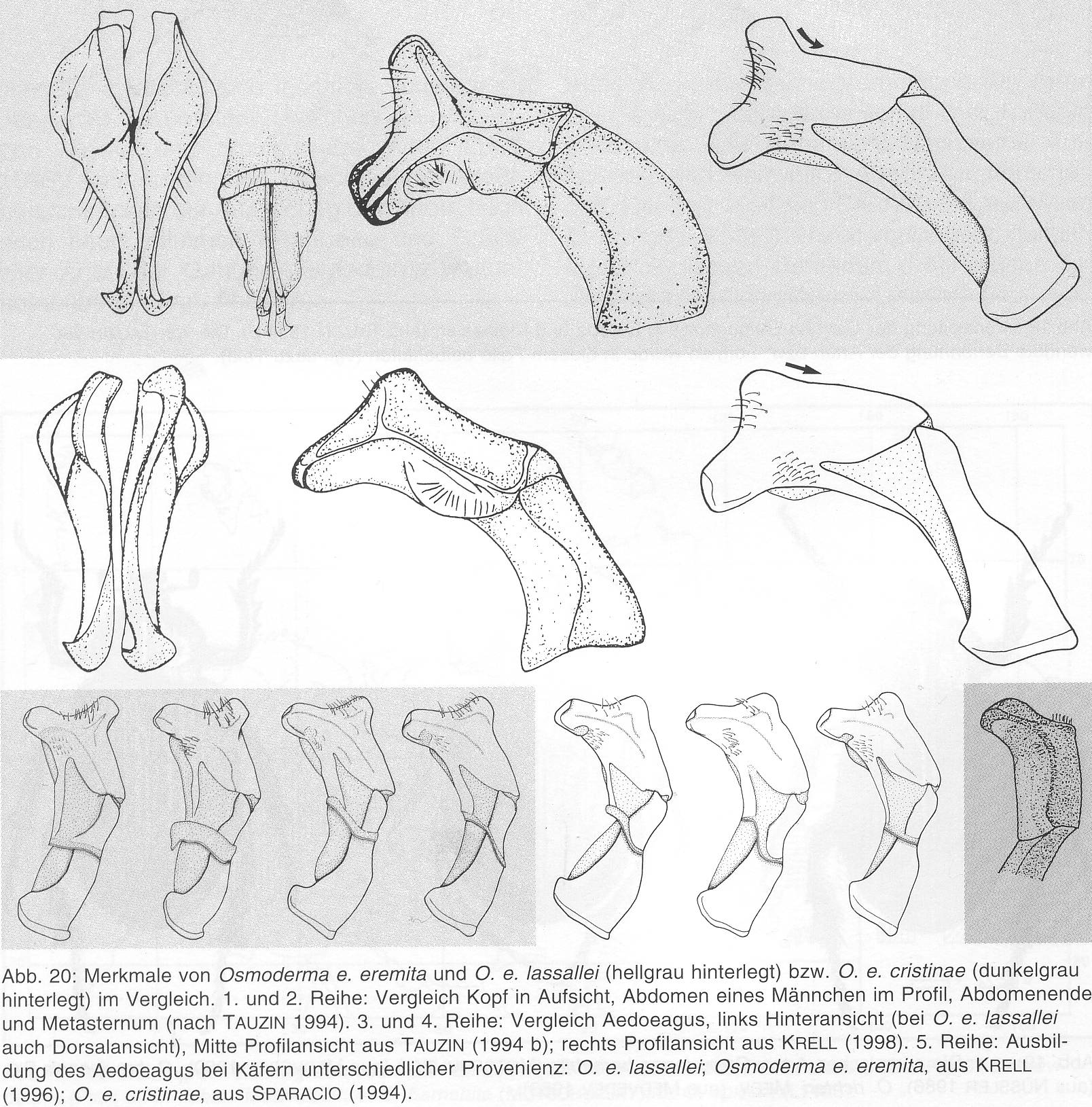 Abb. 08-2: Ausschnitt aus einer Übersicht zum Bau des männlichen Genitals aus: Schaffrath (2003a)
Abb. 08-2: Ausschnitt aus einer Übersicht zum Bau des männlichen Genitals aus: Schaffrath (2003a)
2. Eigene Ergebnisse
Nachfolgende Abbildung zeigt die Variationsbreite der männlichen Genitalform des Eremiten von verschiedenen Fundorten in Sachsen (Abb. 08-3). Bei Betrachtung der Form der Genitalspitze = Parameren (rechte „Beule“) sind in der Profilansicht sehr unterschiedliche Ausprägungen sichtbar. Vor allem bezogen auf die in Abb. 08-2 mit Pfeilen hervorgehobene Ausprägung der Paramerenspitze gibt es Übergänge. Am deutlichsten ausgeprägt ist diese „Beule“ im rechtselbischen Vorkommen im Dresdner Großen Garten und im linkselbischen Vorkommen bei der Ortschaft Priesa, während diese Beule bei den rechtselbischen Vorkommen bei Karpfenschänke und den linlselbischen Vorkommen bei Grumbach und Mehren sehr flach verläuft. Alle anderen Vorkommen nehmen mehr oder weniger eine Zwischenstellung ein. Geographisch lässt sich somit im Kerngebiet der sächsischen Eremitenpopulationen keine eindeutige Trennlinie ziehen.
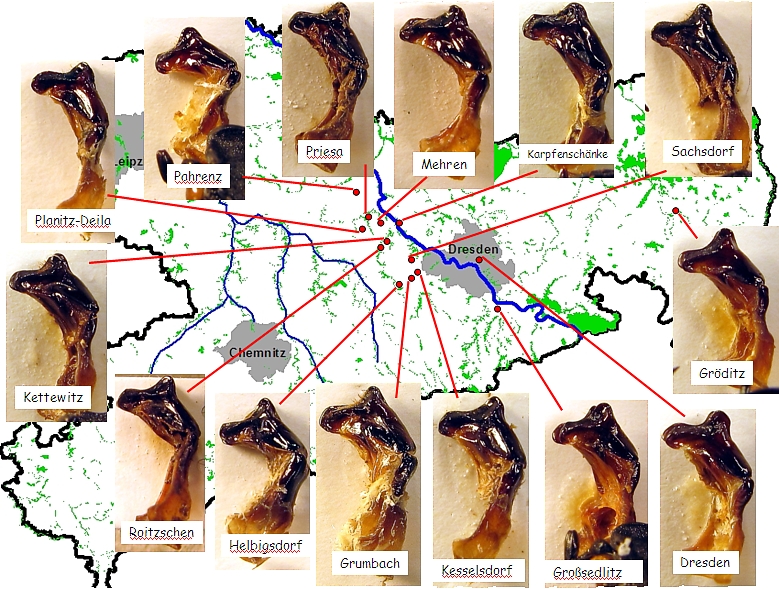 Abb. 08-3: Übersicht zum Bau des männlichen Genitals aus einigen Vorkommen in Sachsen
Abb. 08-3: Übersicht zum Bau des männlichen Genitals aus einigen Vorkommen in Sachsen
3. Diskussion
Falls oben genannte Wiederbesiedlungshypothese tatsächlich der Realität entspricht, wäre es möglich, dass es in Sachsen und speziell im Raum Dresden zum „Zusammentreffen“ der aus Südwest- und Südosteuropa stammenden Eremiten-Populationen gekommen ist, sodass hier eine Vermischung bzw. Überlappung der nacheiszeitlich zugewanderten (Meta?)populationen vorliegt. Unklar ist, ob die Isolation während der letzten Eiszeit zur einer solch genetischen Differenzierung geführt hat, dass sich verschiedene Arten ausgebildet haben oder ob es südlich der Alpen ein zusammenhängendes Verbreitungsareal gab.
Eine ökologische Differenzierung bzw. die Bevorzugung einer bestimmten Baumart scheint es offensichtlich nicht zu geben. Entscheidend für die Besiedlung dürfte die Höhlenbaum-Biotoptradition und das Vorhandensein von ausreichend großen Mulmhöhlen mit einer bestimmten Feuchte und holzzersetzenden Pilze zu sein.
Im Dresdner Raum werden vom Eremiten auch nichtheimische Baumarten besiedelt, wie z.B. Robinie (Robinia pseudoacacia), Roteiche (Quercus rubra), Platane (Platanus sp.), Japanischer Schnurrbaum (Styphnolobium japonicum), Tulpenbaum (Liriodendrum tulipifera) usw. Eine Übersicht über die Baumartenverteilung der Eremitenbrutbäume im Hauptvorkommensgebiet Oberes Elbtal ist in Lorenz (2013) graphisch dargestellt. Etwa 60 % der 1215 kartierten Brutbäume sind Obstbäume, v.a. Kirsche, Apfel und Birne, gefolgt von Eichen (18 %), Linden (8 %) und Kopfweiden (6%).
Im Labor lassen sich Eremitenlarven nicht nur im Mulmsubstrat züchten sondern auch sehr erfolgreich mit Hilfe von Eichen- und Buchenlaub (mündl. Mitteilung M. Petzold).
Eine Möglichkeit zur Klärung des Artstatus im klassischen Sinne wären Zuchten und Kreuzungsversuche, v.a. um zu prüfen, ob die Nachkommen fertil sind oder nicht. Aber auch dies hätte sein Grenzen, wenn es sich tatsächlich um Schwesternarten handelt. Molekulargenetisch kann man sehr wohl feststellen, ob es zwischen zwei Populationen Genfluss gibt oder nicht, und besteht dieser nicht, lässt sich das meist auch morphometrisch nachweisen.
4. Literatur
- Audisio, P., Brustel, H., Carpaneto, G.M., Coletti, G., Mancini, E., Piattella, Trizzino, M., Dutto, M., Antonini, G. & A. DeBiase 2007: Updating the taxonomie and distribution of the european Osmoderma, and strategies for their conservation. – Fragmenta entomologica, Roma, 39 (2): 273-290.-
- Gusakov, A.A. (2002): Scarabaeid beetles of the subfamily Trichinae (Coleoptera, Scarabaeidae) in the Fauna of Russia. Species of the genus Gymnodus Kirby, 1827. – Eidos, Kaluga, 55pp.-
- Hewitt, G.M. (1999): Post-glacial re-colonization of European biota. – Biological Journal of the Linnean Society, 68: 87-112.-
- Krell, F.-T. 1996: Zu Taxonomie, Chorologie und Eidonomie einiger westpaläarktischer Lamellicornia (Coleoptera). – Entomologische Nachrichten und Berichte 40 (4): 217-229.-
- Larsson, C.M., Hedin, J., Svensson, G.P., Tolasch, T. & W. Franke 2003: Charateristic odor of Osmoderma eremita identified as a male-released Pheromone. – Journal of -
- Lorenz, J. (2013): Historische Nachweise, gegenwärtige und Prognose der zukünftigen Bestandssituation des Eremiten (Osmoderma eremita (Scopoli, 1763)) in Sachsen (Coleoptera: Scarabaeidae). – Sächsische Entomologische Zeitschrift 7 (2013/2013): 3-29.-
- Ranius, Th. & J. Hedin 2001: The dispersal rate of a beetle, Osmoderma eremita, living in tree hollows. – Oecologia 126 (3): 363-370.-
- Ranius, Th., Aguado, L.O., Antonsson, K., Ballerio, A., Carpaneto, G.M., Chobot, K., Gjurašin, B., Hanssen, O., Huijbregts, H., Lakatos, F., Martin, O., Neculiseanu, Z., Nikitsky, N.B., Paill, W., Pirnat, A., Rizun, V., Ruicaňescu, A., Stegner, J., Suda, I., Szwałko, P., Tamutis, V., Telnov, D., Tsinkevich, V., Versteirt, V., Vignon, V., Vogeli, M., & P. Zach (2005): Osmoderma eremita (Coleoptera, Scarabaeidae, Cetoniinae) in Europe. – Animal Biodiversity Conservation, 28(1): 1-44.-
- Rössner, E. 2012: Die Hirschkäfer und Blatthornkäfer Ostdeutschlands (Coleoptera: Scarabaeoidea). – Hrsg.: Verein der Freunde & Förderer des Naturkundemuseums Erfurt e. V. 508 S.-
- Schaffrath, U. 2003a: Zu Lebensweise, Verbreitung und Gefährdung von Osmoderma eremita (Scopoli, 1763) (Coleoptera, Scarabaeoidea, Cetoniidae, Trichiinae), Teil 1. - Philippia – Abhandlungen und Berichte aus dem Naturkundemuseum im Ottoneum zu Kassel 3: 157-248.-
- Schaffrath, U. 2003b: Zu Lebensweise, Verbreitung und Gefährdung von Osmoderma eremita (Scopoli, 1763) (Coleoptera, Scarabaeoidea, Cetoniidae, Trichiinae), Teil 2. - Philippia - Abhandlungen und Berichte aus dem Naturkundemuseum im Ottoneum zu Kassel 4: 249-336.-
- Stegner, J. 2002: Der Eremit, Osmoderma eremita (Scopoli, 1763) (Col., Scarabaeidae), in Sachsen: Anforderungen an Schutzmaßnahmen für eine prioritäre Art der FFH-Richtlinie. - Entomologische Nachrichten und Berichte 46: 213-238.-
- Stegner, J. 2004: Bewertungsschema für den Erhaltungszustand von Populationen des Eremiten Osmoderma eremita (Scopoli, 1763). - Naturschutz und Landschaftsplanung 36 (9): 270-276.-
- Stegner, J. & Strzelczyk, P. 2006: Der Juchtenkäfer (Osmoderma eremita), eine prioritäre Art der FFH-Richtlinie. Handreichung für Naturschutz und Landschaftsplanung - VIDUSMEDIA 1. Auflage: 1-43.-
- Tauzin, P. 1994a: Le genre Osmoderma Le Peletier et Audinet-Serville 1828 (Coleoptera, Cetoniidae, Trichiinae, Osmoderatini). Systemique, Biologie et Distibution (I. Partie). – L’Entomologiste 50 (3): 195-214. Paris.-
- Tauzin 1994b: Le genre Osmoderma Le Peletier et Audinet-Serville 1828 (Coleoptera, Cetoniidae, Trichiinae, Osmoderatini). Systemique, Biologie et Distibution (II. Partie). – L’Entomologiste 50 (3): 217-242. Paris.