Die Holz- und Pilzkäferfauna im NSG „Seußlitzer Grund“ nordwestlich von Meißen
1. Vorbemerkungen
Das NSG „Seußlitzer Grund“ umfasst 148,43 ha und liegt östlich der Ortschaft Diesbar-Seußlitz (SMUL 2008). Nachdem das Schutzgebiet 1961 ausgewiesen wurde, erfolgten Anfang der 1990er Jahre im Rahmen der Neuausweisung und der Erarbeitung eines Pflege- und Entwicklungskonzepts sporadische Erfassungen zu Pflanzen- und Tierwelt, u.a. auch zur Holz- und Pilzkäferfauna (LORENZ 1994). Entsprechend einer vorgegebenen Methodik fanden damals wenige Sammelexkursionen statt, wobei in erster Linie stichprobenartige Handfänge durchgeführt wurden, die über das ganze NSG verteilt waren. Das Erfassungsergebnis betrug damals 118 xylobionte Käferarten was im Vergleich zu ähnlichen Untersuchungen in anderen Schutzgebieten als überdurchschnittlich gut bewertet werden konnte. Damals wurden einige faunistisch bemerkenswerte und naturschutzfachlich bedeutsame Arten nachgewiesen, vorwiegend im zentralen Teil des NSG. Es stellte sich die Frage, ob nach über 20 Jahren der Schutzzweck noch gegeben und ein ähnliches Artenspektrum noch vorhanden ist.
Einjährige, meist sporadische Erfassungen sind bei vielen ökofaunistischen Untersuchungen leider die Regel geworden, wenngleich bekannt ist, dass die Ergebnisse meist nicht repräsentativ sind, zumindest nicht für die meisten Insektengruppen sowie bei solch heterogenen, großen Untersuchungsgebieten. Auf Grund ihrer versteckten, hochspezifischen, meist dämmerungs- und nachtaktiven Lebensweise, einer niedrigen Individuenzahl und geringer Vagilität, was den Nachweis besonders schwierig macht, sowie der meist ausgesprochen lokaler, punktuellen Verbreitung können viele xylobionte Käferarten oft erst nach mehrjährigen Erfassungen nachgewiesen werden. Meist sind es aber gerade diese stenöken Spezialisten, die die Beurteilung des naturschutzfachlichen Wertes eines Schutzgebiets implementieren. Als so genannte „hotspots“ für Holz- und Pilzkäfer gelten alte, anbrüchige Höhlenbäume in der Absterbephase. Diese Sonderstrukturen müssen zum richtigen Zeitpunkt gefunden und mit den entsprechenden Methoden untersucht werden, um mit gewisser Wahrscheinlichkeit die tatsächlich wertgebenden Arten, z.B. Urwald-Reliktarten (MÜLLER et al., 2005), nachweisen zu können.
Das Artenspektrum der xylobionten Käfer i.w.S. umfasst in Sachsen ca. 20% der aktuell nachgewiesenen Arten, d.h. etwa 900 Spezies aus fast allen der 100 Käferfamilien. Neben den „echten“ Holzbesiedlern gibt es auch Arten, die sich an Holzpilzen, unter der Rinde oder in Baumhöhlen bzw. im Mulm entwickeln, sodass diese heterogene Gruppe folgendermaßen definiert werden kann:
Zur Gruppe der Holz- und Pilzkäfer zählen alle Arten, deren Entwicklung in lebenden, absterbenden und toten Bäumen erfolgt bzw. die als Larven und (oder) Imagines an oder in Holz, Baumhöhlen bzw. Mulm, Baumpilzen, ausfließendem Baumsaft, Nestern baumhöhlenbrütender Vogelarten sowie in oder unter der Rinde der Bäume leben und direkt oder indirekt an diese Strukturen gebunden sind, einschließlich räuberischer Käferarten, regelmäßig im Holz bzw. unter der Rinde überwinternder Arten sowie Arten, die als „Gäste“ bei holz- und rindennistenden Hymenopteren (z.B. Ameisen, Hornissen, Wespen und Hummeln) leben.
Diese umfangreiche ökologische Gruppe besitzt vielfältige Beziehungen im Nahrungskettengefüge und Stoffkreislauf der Natur. Beispielsweise kommt es durch die Fraßtätigkeit der „echten“ (xylobionten) Holzkäfer zur Oberflächenvergrößerung (Bohrlöcher, Nagespäne) des Holzes, was die Zersetzungsprozesse durch Pilze und Mikroorganismen beschleunigt und somit die Humifizierung und Bodenbildung fördert. Außerdem können die im Holz oder unter der Rinde lebenden Larven einen erheblichen Anteil der Nahrung von Specht, Kleiber, Baumläufer ausmachen und die Imagines gehören zum Beutespektrum von weiteren waldbewohnenden Vogelarten sowie Kleinsäugern (Spitzmäuse, Fledermäuse, Igel usw.). Von großer Bedeutung ist weiterhin die Bereitstellung von Lebensräumen in Form von Bohrlöchern, in denen sich holznistende Grabwespen- und Wildbienenarten ansiedeln, die wiederum von meist artspezifischen Schlupfwespenarten parasitiert werden. Eine relativ große Zahl von Arten dieser Käfergruppe ist dabei außerordentlich stenök, d.h. sie besitzen einen hohen Spezialisierungsgrad, eine geringe ökologische Amplitude, meist ein geringes Ausbreitungsvermögen, durchlaufen einen mehrjährigen Entwicklungszyklus und treten in relativ kleinen Populationen auf. Viele Arten sind an bestimmte Baumarten und besonders stark dimensionierte Exemplare gebunden und stellen bestimmte Ansprüche an Zersetzungsgrad, Feuchtigkeit, Temperatur, Exposition und Pilzbesatz des Holzes. Weiterhin gibt es viele Arten, die mit anderen vergesellschaftet sind (wird eine Art ausgerottet, sterben eine Reihe weiterer Art ebenfalls aus). Außerdem benötigen viele Arten einen Biotopkomplex (sogenannte Tierarten mit integrierenden Raumanspruch, vgl. auch RÜCKRIEM & ROSCHER 1999). Neben dem toten oder absterbenden Baum als Entwicklungshabitat sind blütenreiche Strukturen auf naturnahen Waldwiesen, an Waldrändern und entlang von Wegen und Straßen zur Geschlechterfindung und damit zur Fortpflanzung besonders wichtig, vor allem für Pracht-, Blatthorn- und Bockkäferarten. Insofern kommt alt- und totholzbewohnenden Käfer i.w.S. eine besondere Bedeutung als Qualitätsparameter für Wälder zu (RÜCKRIEM & ROSCHER 1999).
Aufgrund der intensiven forstlichen Bewirtschaftung sind in den letzten Jahrzehnten Alt- und Totholzstrukturen, vor allem stehende, hohle und morsche Bäume größerer Dimensionen, fast flächendeckend beseitigt worden. In den Forsten erreichen die Bäume nur noch ein hiebsreifes Alter von 90-250 Jahren. Damit wird das natürliche, altersbedingte Absterbestadium, in dem es zu einer großen Artenakkumulation kommt, verhindert. Die Folge war und ist, besonders in den Wirtschaftswäldern, ein rapider Rückgang einer Vielzahl von Tier- und Pilzarten, denen alte Höhlenbäume und stärkeres Totholz die einzige Entwicklungsmöglichkeit bietet. In der Käferfauna am stärksten betroffen sind viele Arten aus folgenden Familien: Hirschkäfer (Lucanidae), Bockkäfer (Cerambycidae), Prachtkäfer (Buprestidae), Blatthornkäfer (Scarabaeidae), Schwarzkäfer (Tenebrionidae), Schnellkäfer (Elateridae), Düsterkäfer (Melandryidae) u.a. Die Beseitigung von Alt- und Totholz führte auch zu einem starken Rückgang des Baumpilzbesatzes in den Wäldern, was eine extreme Gefährdung der an diese Pilze gebundenen Käferfauna zu Folge hatte. Viele Vertreter der Familie Pilzkäfer (Erotylidae), Schwammkäfer (Cisidae), Baumschwammkäfer (Mycetophagidae) u.a. sind ebenfalls zunehmend gefährdet.
Die historischen, anthropogenen Einflüsse auf den Wald sind von entscheidender Bedeutung für viele Arten. Je ursprünglicher und in der Vergangenheit unberührter der Wald war, desto wahrscheinlicher ist das Vorhandensein von stenöken Arten, die aus bioindikatorischer Sicht am aussagekräftigsten sind. Eine wesentliche Voraussetzung für das Vorkommen vieler dieser seltenen Holzkäferarten - sogenannte „Reliktarten“ - ist deshalb die Urständigkeit bzw. Biotoptradition, das heißt, dass in den letzten Jahrhunderten durchgängig der gleiche Wald oder zumindest die gleichen einheimischen Baumarten kontinuierlich bis zur Zerfallsphase gestockt haben müssen. Während viele Kennarten der Bodenflora, nach der die jeweilige natürliche Waldgesellschaft charakterisiert wird, oft noch in künstlich aufgeforsteten Beständen vorkommen, da die Samen viele Jahre im Boden überdauern können, führte das Abholzen des ursprünglichen Baumbestandes zum lokalen Aussterben stenöker Käferarten. Eine Ausbreitung der meist ziemlich standortsgebundenen Tiere ist kaum möglich, da ein relativ dichtes Netz von Totholzstrukturen, wie es im „Urwald“ vorhanden ist, in unseren Wirtschaftswäldern fehlt. In Schutzgebieten fristen sie meist ein isoliertes Dasein.
Das Vorkommen einer relativ großen Zahl bestimmter, sowohl an Alt- und Totholz als auch an Baumpilze gebundener Käferarten charakterisiert die Naturnähe eines Waldes besser bzw. dessen naturschützerischen Wert somit wesentlich aussagefähiger als beispielsweise das Vorkommen bestimmter Spechtarten, der in kurzer Zeit größere Entfernungen fliegend zurücklegen können.
2. Methodik
Wie Anfang der 1990er Jahre fanden wiederum mehrere Sammelexkursionen statt bei denen v.a. Kescher- und Klopfschirm sowie Käfersieb zum Einsatz kamen sowie zwei Lichtfänge. Weil sich dies seit geraumer Zeit als effektive Erfassungsmethode bewährt hat, kamen zusätzlich sogenannte Lufteklektoren (Baumkronen-Fensterkreuzfallen) zum Einsatz. Im Jahr 2015 wurden 4 Lufteklektoren im Blatterslebener Grund aufgehängt, einem nordöstlichen Seitentälchen des oberen Seußlitzgrundes, und in den Jahren 2016 sowie 2017 sind jeweils 5 Lufteklektoren im mittleren bzw. unteren, westlichen Teil des NSG von April bis Oktober aufgebaut worden, wobei sie monatlich geleert wurden (Abb. 1, 2, 3).

Abb. 1: NSG Seußlitzer Grund, Position der Lufteklektoren (2015: gelbe Kästchen; 2016: rote Kästchen; 2017: blaue Kästchen) und Flächen außerhalb des Waldes, auf denen vereinzelt Kescher- und Klopfschirmfänge stattfanden (rot schraffiert)

Abb. 2: Beispiel für die Fensterfalle (Lufteklektor) an Baum-Nr. 8: Der Lufteklektor wurde unmittelbar an der großer Stammhöhle der Rot-Buche befestigt, sodass viele Mulmhöhlenarten nachgewiesen werden konnten
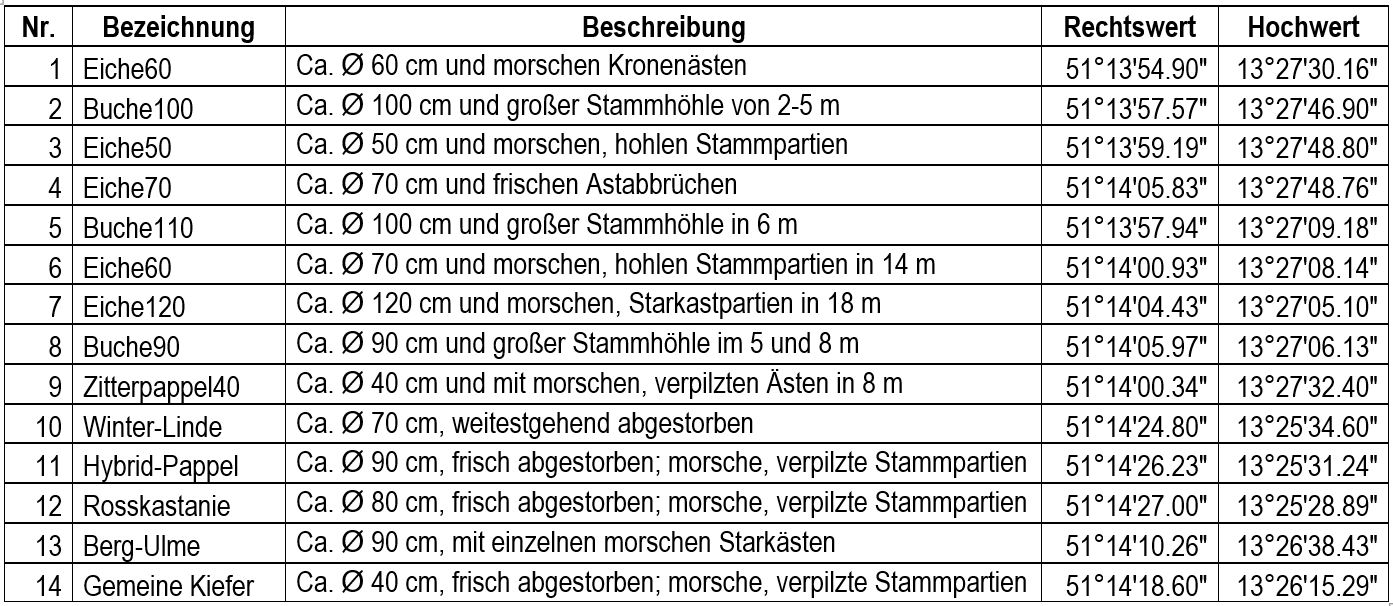
Abb. 3: Liste mit Angaben zu den Lufteklektor-Bäumen
Die Untersuchungen erfolgten mit Unterstützung des Kollegen Olaf Jäger vom Senckenberg Museum für Tierkunde Dresden. Im Herbst 2016 sowie im Jahr 2017 wurde im oberen Seußlitzgrund von Dr. Andre Reimann vom Senckenberg Museum für Tierkunde Dresden eine neue Fangapparatur getestet, eine so genannte Baumkronen-Malaisefalle, die vor allem zur Erfassung bestimmter Dipteren zum Einsatz kam.
3. Ergebnisse xylobionte Käfer
Im Jahr 2015 konnten 385 Käferarten nachgewiesen werden, darunter 183 Holz- und Pilzkäferarten. Beim Lichtfang am 7.7.2015 wurden 108 Käferarten gefunden und in den Lufteklektoren zwischen 67 und 107 Arten je Falle. Im Jahr 2016 konnten mit 366 Käferarten eine ähnlich hohe Zahl nachgewiesen werden wie 2015, wobei sich die Gesamtartenzahl auf 576 erhöht, darunter, wie ein Jahr zuvor etwa 180 Holz- und Pilzkäferarten. Insgesamt erhöht sich die Artenzahl an xylobionten Käfern auf 260, da 80 neue Arten hinzugekommen sind, d.h. sie konnten im Jahr 2015 nicht gefunden werden. Bei den Kescher- und Klopfschirmfängen und sonstigen gelegentlichen Beifängen kamen 2016 knapp 180 Käferarten zusammen. Im Jahr 2017 sind 454 Käferarten nachgewiesen worden, darunter 196 Arten, die in den beiden vorhergehenden Jahren noch nicht gefunden wurden. Beim Lichtfang am 31.7.2017 wurden 77 Käferarten angelockt. Damit erhöht sich die Gesamtzahl nach 3 Jahren Erfassung auf 774 Käferarten, darunter 292 xylobionte Käferarten i.w.S.
Die 292 Holz- und Pilzkäferarten sind in angehängter Tabelle mit Angaben zu Häufigkeit, Gefährdung, Schutzstatus und Ökologie aufgelistet. Laut der bundesdeutschen Roten Liste (GEISER 1998) konnten 10 Arten gefunden werden, die als „vom Aussterben bedroht“ gelten und 43 Arten mit dem Gefährdungsgrad: „stark gefährdet“ sowie 55 Arten, die als „gefährdet“ eingestuft sind. Insgesamt gelang somit der Nachweis von 108 Rote-Liste-Arten entsprechend der bundesdeutschen Roten Liste (GEISER 1998). Nach der Bundesartenschutzverordnung sind 36 Arten besonders geschützt. Etwa 100 Arten und damit ein Drittel des Artenspektrums kann als selten bzw. sehr selten eingestuft werden (siehe Tabelle Spalte „BV“). Außerdem sind nach SCHMIDL & BUSSLER (2004) 31 Arten als sogenannte „Indikatorarten“ eingestuft, d.h. sie besitzen eine große ökologische und bioindikatorische Relevanz und unterstreichen den hohen naturschutzfachlichen Wert des Waldgebiets (Tabelle Spalte „IA“). Bemerkenswert ist der Nachweis von fünf sogenannten „Urwald-Reliktarten“ (nach MÜLLER et al. 2005, ECKELT 2017, LORENZ 2010) (Tabelle Spalte „UR“).
Entsprechend der Ökologie besitzen Altholz- und Frischholzbesiedler mit 107 bzw. 97 Arten etwa gleich große Anteile, gefolgt von Holzpilzbesiedlern mit 54 Arten und Arten mit xylobionten Sonderbiologieen mit 21 Arten sowie 14 Mulmhöhlenbesiedlern (Tabelle Spalte „ÖG“).
Einige faunistische Besonderheiten bzw. naturschutzfach besonders relevante Arten, die gesetzlich geschützt und/oder einen hohen Gefährdungsgrad besitzen, sind beispielsweise:
Osmoderma eremita
Da der Eremit oder Juchtenkäfer als prioritäre Art der FFH-Richtlinie den höchsten Schutzstatus in Europa genießt, sind sowohl die Käfer und Larven als auch seine Brutbäume grundsätzlich geschützt. Es gibt also ausdrücklich ein Tötungsverbot für die Tiere und zusätzlich ein Zerstörungsverbot seiner Entwicklungsstätten. Die Art ist an alte, mit Mulmhöhlen ausgestattete Laubbäume gebunden und wurde sowohl am Ostrand des UG, d.h. im oberen Blatterslebener Grund bzw. in einer hohlen Kirsche in einem Streuobstwiesenrest als auch im unteren Seußlitzgrund bzw. in einer hohlen Rosskastanie nachgewiesen.

Abb. 4: Juchtenkäfer (Osmoderma eremita) (Körperlänge: ca. 3,5 cm)
Die für die Besiedlung des Juchtenkäfers geeignete Baumhöhle muss ein spezifisches, relativ konstantes Innenklima aufweisen, nicht allzu viel Feuchtigkeit aufnehmen, und das Holz ist von bestimmten phytopathogenen bzw. saprophytischen Pilzen bereits teilweise aufgeschlossen, sodass sich erdfeuchter Mulm gebildet hat. Auf Grund ihres geringen Ausbreitungsvermögens zeugen Vorkommen des Juchtenkäfers von einer großen Faunentradition, d.h. auf gleichem Standort haben sich in den letzten Jahrhunderten ähnliche Habitatstrukturen befunden: alte, anbrüchige Laubbäume mit Baumhöhlen. Der Juchtenkäfer gilt sowohl nach der sächsischen als auch nach der bundesdeutschen Roten Liste als „stark gefährdet“. Entsprechend des europäischen Schutzgebietssystems NATURA 2000 bzw. der FFH-Richtlinie der EU handelt es sich bei Osmoderma eremita um eine „prioritäre Art“ des Anhanges II (Tier- und Pflanzenarten von gemeinschaftlichem Interesse, für deren Erhaltung besondere Schutzgebiete ausgewiesen werden müssen) und um eine Art des Anhanges IV (Streng zu schützende Tier- und Pflanzenarten von gemeinschaftlichem Interesse).
Die Vorkommen des Juchtenkäfers (Osmoderma eremita) konzentrieren sich vor allem in wärmebegünstigten Lagen der großen Flusstäler und des Hügellandes sowie der Ebene. Der Hauptverbreitungsschwerpunkt befindet sich im Bereich des Elbtales zwischen Pirna, Dresden, Meißen und Riesa sowie Döbeln und Leisnig. Nach bisherigem Kenntnisstand gehört das UG zum nördlichen Rand des derzeit bekannten Areals in Sachsen. In den umliegenden Streuobstwiesen gibt es weitere Brutbäume, aber nur wenige Kilometer nördlich und östlich in der intensiv agrarindustriell genutzten Großenhainer Pflege sind keine Fundorte mehr bekannt.
Der Hirschkäfer (Lucanus cervus) ist eine weitere FFH-Käferart, die im Gebiet vorkommt, jedoch im Rahmen dieser Untersuchung nicht gefunden wurde. Es gibt aber Nachweise aus den Jahren 2007 und 2012, die im Rahmen des FFH-Artmonitorings dokumentiert werden konnten.
Protaetia fieberi
Diese sehr seltene Goldkäferart entwickelt sich in Baumhöhlen mit Mulm. Die Larven stellen ähnlich hohe spezifische Anforderungen an das Mulmsubstrat, wie der Eremit. Im Gegensatz zum Marmorierten Goldkäfer hat diese Art ein sehr begrenztes Verbreitungsareal in Sachsen, denn nur in der Umgebung um Meißen sind Funde bekannt, beispielsweise im Saubachtal bei Gauernitz.

Abb. 5: Fieber‘s Goldkäfer (Protaetia fieberi) (2,5 cm)
Protaetia marmorata
Der Marmorierte Goldkäfer entwickelt sich manchmal zusammen mit dem Eremiten in der gleichen Baumhöhle. Die Larven stellen ähnliche, aber nicht ganz so spezifische Anforderungen an das Mulmsubstrat. Insofern kann die Art als „Vorbesiedler“ bzw. „Mulmhöhlenbereiter“ für den Eremiten dienen, weil sie kleinere Höhlen besiedelt, die sich dann über Jahre vergrößern, oder sie ist ein Nachbesiedler von ehemals vom Eremiten genutzten Mulmhöhlen.

Abb. 6: Marmorierter Goldkäfer (Protaetia marmorata) (2,5 cm)
Gnorimus nobilis
Der Grüne Edelscharrkäfer entwickelt sich in Baumhöhlen mit Mulm. Die Larven stellen ähnlich hohe spezifische Anforderungen an das Mulmsubstrat, wie der Eremit. Im Gegensatz zum Marmorierten Goldkäfer wird diese Art aber nur selten gefunden, scheint aber auch eher in submontanen Gegenden vorzukommen und enger an Wälder gebunden zu sein, während die drei vorher genannten Arten öfters auch in Obstbäumen in der offenen Landschaft anzutreffen sind.

Abb. 7: Grüner Edelscharrkäfer (Gnorimus nobilis) (ca. 1,5 cm)
Agrilus ater
Vom Pappel-Prachtkäfer gab es aus Sachsen nur einen etwa 100 Jahre zurückliegender Nachweis aus der Leipziger Region. Im Jahr 2010 wurde die Art wiedergefunden (KWAST 2010). Seit ein paar Jahren scheint die Art etwas häufiger zu werden, wahrscheinlich auch wegen des zunehmenden Absterbens der Hybrid-Pappeln, die letzmalig zu DDR-Zeiten in größerem Umfange, v.a. in der Feldflur gepflanzt wurden. Unter der abgelösten Rinde toter Pappeln kann man ziemlich regelmäßig die charakteristischen Fraßgänge finden. Die sehr flüchtigen Käfer werden kaum nachgewiesen, vorwiegend mit Lufteklektoren, wie hier im Seußlitzgrund.

Abb. 8: Pappel-Prachtkäfer (Agrilus ater) (Körperlänge: ca. 1 cm)
Sinodendron cylindricum
Der Kopfhornschröter gehört zu den kleineren Verwandten des Hirschkäfers und entwickelt sich beispielsweise im mäßig feuchten, morschen, bereits von Pilzen etwas aufgeschlossenen Holz am Stammfuß von alten, anbrüchigen Laubbäumen. Zudem konnte ein weiterer Hirschkäfer-Verwandter gefunden werden, der Rehschröter (Platycerus caraboides).

Abb. 9: Kopfhornschröter (Sinodendron cylindricum) (ca. 1,5 cm)
Pedostrangalia revestita
Der Rotgelbe Buchenschmalbock gehört zu den seltenen, heimischen Bockkäferarten. Bisher sind nur wenige Fundorte bekannt, beispielsweise nahe der Ortschaft Koblenz im Oberlausitzer Heide- und Teichgebiet oder bei Grimma. Die Entwicklung findet nicht nur an Buche, sondern auch an anderen Laubbaumarten statt. Nicht auszuschließen ist auch die Hybrid-Pappel als Fraßpflanze, denn die Art wurde im Lufteklektor nachgewiesen, der in dieser Baumart hing.

Abb. 10: Buchenschmalbock (Pedostrangalia revestita) (1,5 cm)
Obrium cantharinum
Der Dunkelbeinige Flachdeckenbock entwickelt sich in Pappelästen und lebt vorwiegend in den Baumkronen. Am Boden wird er kaum gefunden. Es gibt bisher erst wenigen Nachweise in Sachsen.

Abb. 11: Flachdeckenbock (Obrium cantharinum) (1 cm)
Axinopalpis gracilis
Der Messerbock kommt vor allem in den Baumkronen von Eichen vor. Er gilt als sehr selten, konnte aber in den vergangenen Jahren mehrfach gefunden werden, v.a. mit Hilfes des Lichtfanges. Offenbar ist die Art an warmen Hochsommernächten besonders flugaktiv und wird vom Licht angelockt.

Abb. 12: Messerbock (Axinopalpis gracilis) (ca. 1 cm)
Xylotrechus rusticus
Der Graue Espenbock galt in Sachsen früher noch als ziemlich selten, scheint aber in den vergangenen Jahren etwas häufiger zu werden. Wahrscheinlich profitiert die Art ähnlich wie der oben genannte Pappel-Prachtkäfer von den vielen derzeit absterbenden Hybridpappeln. Vielleicht auch Nicht auszuschließen ist auch die Hybrid-Pappel als Fraßpflanze, denn die Art wurde im Lufteklektor nachgewiesen, der in dieser Baumart hing.

Abb. 13: Grauer Espenbock (Xylotrechus rusticus) (ca.1,5 cm)
Plagionotus detritus
Der Hornissenbock galt in Sachsen früher auch als ziemlich selten, scheint aber in den vergangenen Jahren etwas häufiger zu werden. Die Art entwickelt sich v.a. an frisch totem Eichenholz.

Abb. 14: Hornissenbock (Plagionotus detritus) (ca.1,5 cm)
Exocentrus adspersus
Der Weißgefleckte Wimperhornbock entwickelt sich vor allem in recht dünnen, absterbenden oder frisch toten Eichenästen während der ähnlich aussehende Linden-Wimperhornbock (Exocentrus lusitanus) regelmäßig von recht dünnen, absterbenden oder frisch toten Lindenästen geklopft werden kann.

Abb. 15: Wimperhornbock (Exocentrus adspersus) (0,6 cm)
Ropalopus femoratus
Der Mattschwarze Scheibenbock ist auch in erster Linie ein Baumkronenbewohner, v.a. von alten Eichen. Am Boden findet man ihn nur selten. Die Art wurde an der großen, alten Eiche mit Hilfe des Lufteklektors Nr. 7 gefangen.
Abb. 16: Schwarzer Scheibenbock (Ropalopus femoratus) (1 cm)(Foto: Vaclav Hanzlik)
Xylotrechus antilope
Der Zierliche Widderbock kommt vor allem an Eichenästen vor. Die Art wurde in den letzten häufiger nachgewiesen.

Abb. 17: Zierlicher Widderbock (Xylotrechus antilope) (0,6 cm)
Cerambyx scopoli
Der Buchenbock oder Kleine Eichenbock entwickelt sich sowohl in Rot-Buchen und Eichen als auch v.a. in Obstbäumen, besonders Kirschen.

Abb. 18: Buchenbock (Cerambyx scopoli) (2 cm)
Saperda perforata
Der Gefleckte Pappelbock scheint ähnlich wie der Pappelprachtkäfer und andere an Pappeln gebundene Arten in den letzten Jahren etwas häufiger zu werden, weil er sich auch in den zunehmend absterbenden Hybridpappeln entwickelt.

Abb. 19: Gefleckter Pappelbock (Saperda perforata) (1,5 cm)
Neatus picipes
Dieser dem Mehlkäfer änliche Schwarzkäfer, entwickelt sich in Mulmhöhlen und im morschen Holz alter Laubbäume. In der Region um Meißen befindet sich ein Verbreitungsschwerpunkt dieser ziemlich seltenen Art, da sie hier meist zusammen mit dem Juchtenkäfer vorkommt, v.a. in den Streuobstwiesen bzw. alten, hohlen Obstbäumen.

Abb. 20: Schwarzkäfer (Neatus picipes) (1,5 cm)
Aeletes atomarius
Diese sehr kleine und sehr seltene Stutzkäferart lebt in großen Baumhöhlen von sehr alten Laubbäumen, v.a. Buchen. Auf Grund dieser engen Abhängigkeit an Biotoptradition und das Vorkommen in Wäldern mit altem Baumbestand gilt er als sogenannte Urwald-Reliktart. Bisher sind nur wenige Nachweise aus der Oberlausitz bekannt (KLAUSNITZER et al. 2009).

Abb. 21: Buchenmulm-Zwergstutzkäfer (Aeletes atomarius) (1 mm) (www.kerbtiere.de)
Thoracophorus corticinus
Dieser kleine und sehr seltene Urwald-Kurzflüglerkäfer lebt unter der verpilzten Rinde von sehr alten Laubbäumen, v.a. Buchen. Auf Grund dieser engen Abhängigkeit an Biotoptradition und das Vorkommen in Wäldern mit altem Baumbestand gilt er als sogenannte Urwald-Reliktart. Es gibt nur wenige Nachweise aus Sachsen, beispielsweise aus dem Moritzburger Wald und dem Röderauwald bei Skassa westlich von Großenhain.

Abb. 22: Urwald-Kurzflügler (Thoracophorus corticinus) (2 mm)
Enicmus planipennis
Die seltene Art lebt v.a. in den Baumkronen alter Kiefern. Es gibt nur wenige Funde aus Sachsen, beispielsweise aus der Oberlausitz (KLAUSNITZER et al. 2009) und bei Torgau (REIKE et al., 2005) sowie im Mittleren Erzgebirge bei Satzung (JÄGER, LORENZ & REIKE 2013)

Abb. 23: Moderkäfer (Enicmus planipennis) (1,5 mm) (Foto: Piezo)
Magdalis caucasicus
Dieser sehr seltene Rüsselkäfer entwickelt sich an Ulme und dürfte durch das Ulmensterben weiterhin an Lebensraum einbüßen.

Abb. 24: Zahnhals-Knospenstecker (Magdalis caucasicus) (0,6 cm)
Corticeus fasciatus
Der Schwarzkäfer Corticeus fasciatus kommt vorwiegend und der losen Rinde abgestorbener Eichen vor. Die Urwald-Reliktart konnte in den vergangenen 10 Jahren an mehreren Fundorten in Sachsen nachgewiesen werden, meist in Waldbeständen und Parks mit Alt-Eichen.

Abb. 25: Rotbindige Linien-Schwarzkäfer (Corticeus fasciatus) (0,5 cm) (Foto: H. Bouyon)
Nachtrag:
Zwei weitere faunistisch interessante Arten, die nach Müller et al. (2005) zu den Urwald-Reliktarten gehören, wurden nach Abschluss der Erfassungen in den letzten beiden Jahren im NSG „Seußlitzer Grund“ gefunden: Ein Vorkommen des Großen Goldkäfers Protatia speciosissima (Scopoli, 1786) (=Protaetia aeruginosa (Linnaeus, 1767)) hat Herr Maik Petzold im oberen Seußlitzgrund nachgewiesen, und Herr Rüdiger Peschel teilte mit, dass er Neomida haemorrhoidalis (Fabricius, 1787) im unteren Seußlitzgrund an einer Buche mit Baumpilzen gefunden hat (vielen Dank den Beiden für die Informationen).

Abb. 19_26: Vom Großen Goldkäfer Protaetia speciosissima gibt es aus Nordsachsen einige wenige Nachweise. Im Elbtal um Meißen und Dresden ist die Art sehr selten.

Abb. 19_27: Der Schwarzkäfer Neomida haemorrholidalis galt bis vor wenigen Jahren als große Seltenheit, kann aber plötzlich an vielen Stellen, v.a. an Rot-Buchen mit Zunderschwamm-Bewuchs gefunden werden.
4. Vorschläge zu Pflege und Entwicklung
Da es sich um ein Natur-Schutz-Gebiet handelt, sollte der Schutz der Natur auch im Vordergrund stehen, was u.a. eine ungestörte Waldentwicklung und damit auch eine weitere Totholz-Anreicherung, v.a. Stehend-Totholz einschließt. Da jedoch ein Weg durchs Gebiet führt, gilt dort die sogenannte Wegesicherungspflicht. Wenn aus Gründen der Verkehrssicherheit Bäume tatsächlich eine Gefahr darstellen, sollten in Wegnähe zumindest standsichere Hochstubben erhalten bleiben und die Bäume nicht wie üblich durch Wurzelhalsschnitt und Stubbenfräsen komplett beseitigt werden. Fremdländische und invasiv sich ausbreitende Baumarten wie Rot-Eiche und Robinie sollten sukzessive entnommen oder durch Ringeln zum Absterben gebracht werden, besonders dort, wo sie heimische Gehölze bedrängen. Andererseits sollten Eichen ebenso gefördert werden wie die Nachpflanzung von Hochstamm-Obstbäumen auf den Streuobstwiesen. In geringem Umfang könnte eine einzelstammweise Entnahme von hochwertigen Stämmen mit Furnierqualität toleriert werden, wenn die Erlöse für Naturschutzmaßnahmen eingesetzt werden und es zur Verbesserung der Schutzsituation im NSG beiträgt.
Aus naturschutzfachlicher und ökologischer Sicht sollte die Wiesenmahd gestaffelt werden, d.h. nie eine Komplettmahd zu einem Zeitpunkt, sondern ein Wechsel von früher und später Mahd, und es müssen immer ca. 20 % der Fläche ungemäht bleiben, jedoch jedes Jahr ein anderer Bereich. Dies setzt allerdings ein Umdenken bei den Behörden voraus, die meist nach „Schema F“ verfahren und naturschutzfachlich unsinnige Förderrichtlinien umsetzen wollen und die örtlichen Bewirtschafter zwingen, fragwürdige Mahdtermine einzuhalten. Entlang des Weges im Talgrund sollte ein Blühsaum erhalten bleiben.
5. Fazit
Der Nachweis von fast 300 xylobionten Käferarten, was etwa ein Drittel des in Sachsen vorkommenden Artenspektrums entspricht, unterstreicht die große Bedeutung des NSG „Seußlitzer Grund“ als Refugium einer überduchschnittlich hohen Zahl schutzwürdiger xylobionter Arten. Rund 100 der nachgewiesenen Arten stehen auf der bundesdeutschen Roten Liste und 36 Arten sind laut BArtSchV gesetzlich geschützt.
Die dreijährige Erfassung wurde auf rein ehrenamtlicher Basis durchgeführt. Der daraus resultierende umfangreiche Bericht ist der Unteren Naturschutzbehörde Meißen zur Verfügung gestellt worden, die lediglich die Ausnahmegenehmigung zur Erfassung der Insekten erteilte. Leider hat sie es weder für nötig erachtet, den Eingang des Berichts zu bestätigen, noch sich dafür zu bedanken oder eine finanzielle Unterstützung zu gewähren.
6. Literatur
- BÖHME, J. (2001): Phytophage Käfer und ihre Wirtspflanzen in Mitteleuropa. Ein Kompendium. – bioform. 132 S.
- ECKELT, A. und 23 weitere Autoren (2017): “Primeval forest relict beetles” of Central Europe: a set of 168 umbrella species for the protection of primeval forest remnants. - Journal of Insect Conservation https://doi.org/10.1007/s10841-017-0028-6. 14 S.
- FREUDE, H., HARDE, K.W. & G.A. LOHSE (Hrsg.) (1964- 83): Die Käfer Mitteleuropas. Band 1-11. - Verlag Goecke & Evers, Krefeld.
- GEISER, R. (1998): Rote Liste der Käfer (Coleoptera) Deutschlands - Schriftenreihe für Landschaftspflege und Naturschutz, Heft 55: 168-230.
- JÄGER, O., LORENZ, J. & H.-P. REIKE (2013): 3. Treffen ostsächsischer Entomologen. – Mitteilungen Sächsischer Entomologen, Band 32 (Nr. 103): 25-28.
- KÄSTNER, T. (2016): Ein neues Vorkommen der Gemeinen Plumpschrecke Isophya kraussii Brunner von Wattenwyl, 1878 im Landkreis Meißen (Insecta: Orthoptera, Enisfera). - Sächsische Entomologische Zeitschrift, Band 8 (2014/2015): 218-222.
- KOCH, K. (1989a, 1989b, 1992): Die Käfer Mitteleuropas. Ökologie, Bde. 1-3. - Verlag Goecke & Evers, Krefeld.
- KÖHLER, F. (2000): Totholzkäfer in Naturwaldzellen des nördlichen Rheinlands. - Hrsg.: Landesanstalt für Ökologie, Bodenordnung und Forsten / Landesamt für Agrarordnung NRW, LÖBF-Schriftenreihe, Band 18: 351 S.
- KLAUSNITZER, B. (1994): Rote Liste Bockkäfer des Freistaates Sachsen. Ausgabe 1994. - Arbeitsmaterialien Naturschutz: 10 S.
- KLAUSNITZER, B. (1995): Rote Liste Blatthornkäfer und Hirschkäfer des Freistaates Sachsen. Stand 1995. - Materialien zu Naturschutz und Landschaftspflege 5: 10 S.
- KLAUSNITZER, B. (2004): Bemerkungen zur Biologie und Verbreitung einiger Meloidae (Col.) in Mitteleuropa. – Entomologische Nachrichten und Berichte, 48 (3/4): 261-267.
- KLAUSNITZER, B. (2016): Rote Liste und Artenliste Sachsen. Wasserbewohnende Käfer. – Hrsg.: Sächsisches Landesamt für Umwelt, Landwirtschaft und Geologie. 76 S.
- KLAUSNITZER, B., BEHNE, L., FRANKE, R., GEBERT, J., HOFFMANN, W., HORNIG, U., JÄGER, O., RICHTER, W., SIEBER, M. & VOGEL, J. (2009): Die Käferfauna (Coleoptera) der Oberlausitz. Teil 1. – Entomologischen Nachrichten und Berichte, Beiheft 12: 252 S., 1 Karte. Dresden.
- LOHSE, G.A. & W.H. LUCHT (Hrsg.) (1989): Die Käfer Mitteleuropas. Erster Supplementband mit Katalogteil (Bd. 12), Verlag Goecke & Evers, Krefeld.
- LOHSE, G.A. & W.H. LUCHT (Hrsg.) (1992): Die Käfer Mitteleuropas. Zweiter Supplementband mit Katalogteil (Bd. 13), Verlag Goecke & Evers, Krefeld.
- LOHSE, G.A. & W.H. LUCHT (Hrsg.) (1994): Die Käfer Mitteleuropas. Dritter Supplementband mit Katalogteil (Bd. 14), Verlag Goecke & Evers, Krefeld.
- LORENZ, J. (2006): Bedeutung, Gefährdung und Schutz von Alt- und Totholzlebensräumen sowie Ergebnisse mehrjähriger Untersuchungen in Dresden einschließlich landschaftspflegerischer Umsetzung. - NSI - Projektberichte 2/2006 (Hrsg.: AG Naturschutzinstitut Region Dresden e.V.): 2., überarbeitete und erweitere Auflage, 20seitige Broschüre.
- LORENZ, J. (2009): Errichtung von Totholz-Lagerplätzen. – NSI-Projektberichte Praktischer Artenschutz 1/2009 (Hrsg.: AG Naturschutzinstitut Region Dresden e.V.): 2., überarbeitete Auflage, 4seitiges Faltblatt.
- LORENZ, J. (2010): „Urwaldrelikt“-Käferarten in Sachsen (Coleoptera part.). – Sächsische Entomologische Zeitschrift 5: 69-98.
- LORENZ, J. (2012): Totholz stehend lagern – eine sinnvolle Kompensationsmaßnahme? – Naturschutz und Landschaftsplanung 44 (10): 300-306.
- LORENZ, J. (2013): Historische Nachweise, gegenwärtige und Prognose der zukünftigen Bestandssituation des Eremiten (Osmoderma eremita (Scopoli, 1763)) in Sachsen (Coleoptera: Scarabaeidae). – Sächsische Entomologische Zeitschrift 7 (2013/2013): 3-29.
- LUCHT, W.H. & B. KLAUSNITZER (Hrsg.) (1998): Die Käfer Mitteleuropas. Vierter Supplementband. - Verlag Goecke & Evers, Krefeld im G. Fischer Verlag Jena.
- MÜLLER, J., BUßLER, H., BENSE, U., BRUSTEL, H., FLECHTNER, G., FOWLES, A., KAHLEN, M., MÖLLER, G., MÜHLE, H., SCHMIDL, J. & P. ZABRANSKY (2005): Urwaldrelikt-Arten – Xylobionte Käfer als Indikatoren für Strukturqualität und Habitattradition (Insecta, Coleoptera part.). – waldoekologie online, H. 2, S. 106-113.
- REIKE, H.-P., RATSCHKER, U.M., NICOLAI, V. & A. JÄPEL (2005): Moderkäferzönosen (Col., Lathridiidae) in ostdeuschen Kiefernwäldern. – Entomologische Nachrichten und Berichte, Heft 49/3-4: 207-214.
- RÜCKRIEM, CH. & S. ROSCHER (1999): Empfehlungen zur Umsetzung der Berichtspflicht gemäß Artikel 17 der Fauna-Flora-Habitat-Richtlinie: Ergebnisse des Life-Projekts "Beurteilung des Erhaltungszustandes natürlicher Lebensräume gemäß der FFH-Richtlinie" des Bundesamtes für Naturschutz von 1996 – 1998. – Bundesamt für Naturschutz Schriften-Vertrieb im Landwirtschaftsverlag: 456 Seiten.
- SCHMIDL, J. & H. BUßLER (2004): Ökologische Gilden xylobionter Käfer Deutschlands. – Naturschutz und Landschaftsplanung 36, (7): 202-218.
- SMUL (2008): Naturschutzgebiete in Sachsen. – Sächsisches Staatsministerium für Umwelt und Landwirtschaft (Hrsg.). 720 S.
Liste der von 2015 bis 2017 im Seußlitzgrund (einschl. Blatterslebener Grund) nachgewiesenen Holz- und Pilzkäferarten mit Angaben zu Gefährdung, Schutzstatus, Häufigkeit und Ökologie
Artenliste_Seusslitzgrund_Lorenz2020.xls
| Familie | Art | ’15 | ’16 | ’17 | RLDt | RLSn | BV | H | ÖG | IA | UR | BA | BE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Histeridae | Plegaderus caesus | x | x | . | nb | . | sh | A | . | . | LH | s | |
| Histeridae | Abraeus granulum | x | x | 3 | nb | . | s | A | . | . | LH | s | |
| Histeridae | Aeletes atomarius | x | x | 1 | nb | . | ss | xS | ! | 2 | Bu | s | |
| Histeridae | Dendrophilus punctatus | x | x | x | . | nb | . | sh | xS | . | . | Ei | s |
| Histeridae | Paromalus flavicornis | x | x | x | . | nb | . | sh | A | . | . | LH | s |
| Histeridae | Paromalus parallelepipedus | x | . | nb | . | h | F | . | . | NH | s | ||
| Histeridae | Hololepta plana | x | x | . | nb | . | mh | F | . | . | Pa | s/l | |
| Cholevidae | Nemadus colonoides | x | x | 3 | nb | . | mh | xS | . | . | LH | s | |
| Cholevidae | Anemadus strigosus | x | x | 2 | nb | . | s | xS | . | . | Ei | s | |
| Leiodidae | Anisotoma humeralis | x | x | . | nb | . | sh | P | . | . | LH | s/l | |
| Leiodidae | Agathidium nigripenne | x | x | x | . | nb | . | sh | P | . | . | Bu | s/l |
| Scydmaenidae | Scydmaenus perrisi | x | x | 2 | nb | . | s | xS | . | . | LH | s | |
| Staphylinidae | Siagonium quadricorne | x | 3 | nb | . | mh | F | . | . | LH | l | ||
| Staphylinidae | Scaphisoma agaricinum | x | x | . | nb | . | sh | P | . | . | LH | l | |
| Staphylinidae | Thoracophorus corticinus | x | x | x | 1 | nb | . | ss | xS | . | 2 | Bu | s |
| Staphylinidae | Phyllodrepa melanocephala | x | x | 3 | nb | . | s | xS | . | . | LH | s | |
| Staphylinidae | Phyllodrepa nigra | x | x | . | nb | . | ss | xS | . | . | LH | s | |
| Staphylinidae | Dropephylla ioptera | x | x | x | . | nb | . | mh | A | . | . | LH | s/l |
| Staphylinidae | Xylostiba bosnica | x | . | nb | . | ss | F | . | . | LH | s | ||
| Staphylinidae | Hypnogyra angularis | x | x | x | 3 | nb | . | h | A | . | . | LH | s/l |
| Staphylinidae | Hesperus rufipennis | x | x | x | 2 | nb | . | s | A | . | . | Bu | s |
| Staphylinidae | Quedius dilatatus | x | x | x | 3 | nb | . | mh | xS | . | . | LH | s |
| Staphylinidae | Quedius microps | x | 3 | nb | . | ss | M | . | . | LH | s | ||
| Staphylinidae | Quedius brevicornis | x | x | 3 | nb | . | s | xS | . | . | Ei | s | |
| Staphylinidae | Sepedophilus testaceus | x | x | . | nb | . | sh | A | . | . | L/N | s/l | |
| Staphylinidae | Sepedophilus bipunctatus | x | . | nb | . | h | A | . | . | LH | s/l | ||
| Staphylinidae | Oligota granaria | x | . | nb | . | s | A | . | . | Bu | s/l | ||
| Staphylinidae | Gyrophaena manca | x | . | nb | . | sh | P | . | . | LH | s | ||
| Staphylinidae | Cyphea curtula | x | 2 | nb | . | mh | F | . | . | LH | s/l | ||
| Staphylinidae | Placusa pumilio | x | . | nb | . | s | F | . | . | Ul | s/l | ||
| Staphylinidae | Homalota plana | x | x | x | . | nb | . | h | F | . | . | L/N | s/l |
| Staphylinidae | Anomognathus cuspidatus | x | . | nb | . | h | F | . | . | L/N | s/l | ||
| Staphylinidae | Leptusa pulchella | x | . | nb | . | sh | A | . | . | LH | l | ||
| Staphylinidae | Euryusa optabilis | x | . | nb | . | s | xS | . | . | LH | l | ||
| Staphylinidae | Thamiaraea cinnamomea | x | 3 | nb | . | s | xS | . | . | Ei | s | ||
| Staphylinidae | Phloeopora teres | x | . | nb | . | mh | F | . | . | LH | s/l | ||
| Staphylinidae | Phloeopora testacea | x | x | . | nb | . | sh | F | . | . | NH | s | |
| Staphylinidae | Phloeopora corticalis | x | x | x | . | nb | . | sh | F | . | . | LH | s/l |
| Staphylinidae | Phloeopora scribae | x | . | nb | . | s | F | . | . | LH | s/l | ||
| Staphylinidae | Ischnoglossa prolixa | x | . | nb | . | s | F | . | . | L/N | s/l | ||
| Staphylinidae | Bibloporus bicolor bicolor | x | x | x | . | nb | . | sh | A | . | . | LH | l |
| Staphylinidae | Bibloporus minutus | x | . | nb | . | h | A | . | . | LH | s/l | ||
| Staphylinidae | ? Euplectus kirbii kirbii | x | 1 | nb | . | s | A | . | . | LH | s/l | ||
| Staphylinidae | Euplectus piceus | x | . | nb | . | mh | A | . | . | LH | l | ||
| Staphylinidae | Euplectus mutator | x | x | . | nb | . | mh | A | . | . | LH | s/l | |
| Staphylinidae | Plectophloeus fischeri | x | . | nb | . | h | A | . | . | NH | s | ||
| Staphylinidae | Batrisus formicarius | x | . | nb | . | mh | xS | . | . | LH | s | ||
| Staphylinidae | Batrisodes delaporti | x | . | nb | . | mh | xS | . | . | LH | s/l | ||
| Staphylinidae | Batrisodes venustus | x | . | nb | . | s | xS | . | . | LH | s/l | ||
| Staphylinidae | Batrisodes unisexualis | x | 3 | nb | . | s | xS | . | . | LH | s | ||
| Cantharidae | Malthinus flaveolus | x | x | x | . | nb | . | sh | A | . | . | LH | s/l |
| Cantharidae | Malthodes sp. | x | . | . | . | ? | A | . | . | LH | s/l | ||
| Malachiidae | Malachius bipustulatus | x | x | x | . | nb | . | sh | A | . | . | LH | s/l |
| Dasytidae | Aplocnemus impressus | x | . | nb | . | h | A | . | . | NH | s/l | ||
| Dasytidae | Dasytes plumbeus | x | x | . | nb | . | sh | A | . | . | LH | s/l | |
| Dasytidae | Dasytes aeratus | x | x | . | nb | . | h | A | . | . | LH | s | |
| Cleridae | Tillus elongatus | x | x | x | 3 | nb | . | h | A | . | . | LH | s |
| Cleridae | Opilo mollis | x | x | . | nb | . | mh | A | . | . | Bu | s | |
| Cleridae | Thanasimus formicarius | x | . | nb | . | h | F | . | . | NH | s/l | ||
| Lymexylidae | Lymexylon navale | x | 3 | nb | . | mh | A | ! | . | Ei | s | ||
| Elateridae | Ampedus sanguineus | x | x | . | nb | . | h | A | . | . | L/N | s/l | |
| Elateridae | Ampedus pomorum | x | x | x | . | nb | . | sh | A | . | . | L/N | s/l |
| Elateridae | Ampedus nigroflavus | x | x | x | 3 | nb | . | mh | A | . | . | LH | s/l |
| Elateridae | Ampedus elongatulus | x | 3 | nb | . | s | A | . | . | LH | l | ||
| Elateridae | Brachygonus megerlei | x | x | 2 | nb | . | s | A | ! | . | LH | s | |
| Elateridae | Procraerus tibialis | x | x | 2 | nb | . | s | M | ! | . | LH | s | |
| Elateridae | Melanotus villosus | x | x | . | nb | . | sh | A | . | . | LH | l | |
| Elateridae | Melanotus castanipes | x | . | nb | . | h | A | . | . | Bu | l | ||
| Elateridae | Anostirus purpureus | x | . | nb | . | mh | A | . | . | L/N | s/l | ||
| Elateridae | Calambus bipustulatus | x | 3 | nb | . | s | A | . | . | Ei | s | ||
| Elateridae | Denticollis linearis | x | x | . | nb | . | h | A | . | . | L/N | l | |
| Eucnemidae | Melasis buprestoides | x | x | x | . | nb | . | h | F | . | . | LH | s/l |
| Eucnemidae | Isorhipis melasoides | x | 2 | nb | . | ss | F | . | . | LH | s/l | ||
| Eucnemidae | Eucnemis capucina | x | x | x | 3 | nb | . | h | A | . | . | LH | s |
| Eucnemidae | Dromaeolus barnabita | x | x | 2 | nb | . | s | A | ! | . | LH | s | |
| Eucnemidae | Microrhagus lepidus | x | 3 | nb | . | ss | A | . | . | LH | s | ||
| Eucnemidae | Hylis olexai | x | x | x | 3 | nb | . | mh | A | . | . | L/N | s |
| Eucnemidae | Hylis cariniceps | x | 3 | nb | . | s | A | . | . | L/N | s | ||
| Buprestidae | Anthaxia nitidula | x | x | . | nb | § | h | F | . | . | LH | s | |
| Buprestidae | Anthaxia helvetica | x | . | nb | § | mh | F | . | . | Fi | s/l | ||
| Buprestidae | Agrilus ater | x | 2 | nb | § | ss | F | ! | . | Pa | s | ||
| Buprestidae | Agrilus biguttatus | x | . | nb | . | mh | F | . | . | Ei | s | ||
| Buprestidae | Agrilus laticornis | x | . | nb | § | h | F | . | . | Ei | s | ||
| Buprestidae | Agrilus angustulus | x | . | nb | § | h | F | . | . | LH | s | ||
| Buprestidae | Agrilus sulcicollis | x | x | x | . | nb | § | h | F | . | . | Ei | s |
| Scirtidae | Prionocyphon serricornis | x | x | 3 | 2 | . | mh | xS | . | . | LH | s | |
| Dermestidae | Attagenus punctatus | x | 2 | nb | . | mh | xS | ! | . | Bu | s | ||
| Dermestidae | Trinodes hirtus | x | 3 | nb | . | mh | xS | . | . | Ei | s | ||
| Bothrideridae | Bothrideres bipunctatus | x | 1 | nb | . | s | A | ! | . | LH | s | ||
| Cerylonidae | Cerylon histeroides | x | x | x | . | nb | . | sh | A | . | . | LH | s/l |
| Cerylonidae | Cerylon ferrugineum | x | x | x | . | nb | . | sh | A | . | . | LH | s/l |
| Nitidulidae | Epuraea pallescens | x | . | nb | . | mh | F | . | . | LH | l | ||
| Nitidulidae | Epuraea longula | x | . | nb | . | mh | F | . | . | LH | s | ||
| Nitidulidae | Epuraea distincta | x | 3 | nb | . | mh | P | . | . | LH | s/l | ||
| Nitidulidae | Epuraea variegata | x | x | . | nb | . | sh | P | . | . | LH | s/l | |
| Nitidulidae | Glischrochilus quadriguttatus | x | x | . | nb | . | mh | F | . | . | L/N | s/l | |
| Monotomidae | Rhizophagus dispar | x | x | . | nb | . | sh | F | . | . | L/N | s/l | |
| Monotomidae | Rhizophagus bipustulatus | x | x | x | . | nb | . | sh | F | . | . | L/N | s/l |
| Monotomidae | Rhizophagus fenestralis | x | . | nb | . | mh | F | . | . | LH | l | ||
| Silvanidae | Silvanus bidentatus | x | . | nb | . | h | F | . | . | L/N | s/l | ||
| Silvanidae | Silvanus unidentatus | x | x | . | nb | . | h | A | . | . | LH | s/l | |
| Silvanidae | Uleiota planatus | x | x | . | nb | . | sh | A | . | . | L/N | s/l | |
| Erotylidae | Tritoma bipustulata | x | x | x | . | nb | . | sh | P | . | . | LH | s/l |
| Erotylidae | Dacne bipustulata | x | x | x | . | nb | . | sh | P | . | . | LH | s/l |
| Biphyllidae | Diplocoelus fagi | x | x | . | nb | . | h | P | . | . | LH | s/l | |
| Cryptophagidae | Cryptophagus micaceus | x | x | x | 2 | nb | . | h | xS | . | . | LH | s |
| Cryptophagidae | Cryptophagus labilis | x | x | x | 2 | nb | . | mh | M | . | . | LH | s |
| Cryptophagidae | Atomaria turgida | x | . | nb | . | h | A | . | . | Fi | l | ||
| Cryptophagidae | Atomaria pulchra | x | . | nb | . | mh | P | . | . | L/N | l | ||
| Laemophloeidae | Laemophloeus monilis | x | 3 | nb | . | s | F | . | . | Bu | s/l | ||
| Laemophloeidae | Placonotus testaceus | x | . | nb | . | s | F | . | . | LH | l | ||
| Laemophloeidae | Cryptolestes duplicatus | x | x | . | nb | . | mh | F | . | . | Ei | s/l | |
| Laemophloeidae | Leptophloeus alternans | x | . | nb | . | h | F | . | . | NH | l | ||
| Laemophloeidae | Lathropus sepicola | x | 2 | nb | . | s | F | . | . | LH | s | ||
| Latridiidae | Latridius hirtus | x | x | x | 3 | nb | . | mh | P | . | . | Ei | s |
| Latridiidae | Enicmus brevicornis | x | x | x | 3 | nb | . | s | P | . | . | LH | s/l |
| Latridiidae | Enicmus fungicola | x | x | . | nb | . | h | P | . | . | LH | l | |
| Latridiidae | Enicmus planipennis | x | 1 | nb | . | ss | P | . | . | NH | l | ||
| Latridiidae | Enicmus atriceps | x | x | 2 | nb | . | ss | P | . | . | LH | s/l | |
| Latridiidae | Corticaria alleni | x | 2 | nb | . | ss | A | . | . | Hbu | s | ||
| Latridiidae | Corticarina parvula | x | 2 | nb | . | h | P | . | . | NH | s/l | ||
| Mycetophagidae | Litargus connexus | x | x | . | nb | . | sh | P | . | . | LH | s/l | |
| Mycetophagidae | Mycetophagus quadripustulatus | x | x | x | . | nb | . | h | P | . | . | LH | s/l |
| Mycetophagidae | Mycetophagus piceus | x | x | 3 | nb | . | h | P | . | . | LH | s/l | |
| Mycetophagidae | Mycetophagus salicis | x | 2 | nb | . | mh | P | . | . | LH | s/l | ||
| Mycetophagidae | Mycetophagus atomarius | x | . | nb | . | h | P | . | . | LH | s/l | ||
| Mycetophagidae | Mycetophagus quadriguttatus | x | x | . | nb | . | mh | P | . | . | Ei | s | |
| Mycetophagidae | Mycetophagus fulvicollis | x | x | x | 2 | nb | . | mh | P | ! | . | LH | s |
| Mycetophagidae | Mycetophagus populi | x | x | x | 2 | nb | . | s | P | ! | . | LH | s |
| Colydiidae | Pycnomerus terebrans | x | x | x | 1 | nb | . | mh | A | ! | . | LH | s |
| Colydiidae | Synchita mediolanensis | x | 1 | nb | . | E17 | A | . | . | LH | s | ||
| Colydiidae | Synchita undata | x | x | 3 | nb | . | mh | P | ! | . | LH | s/l | |
| Colydiidae | Bitoma crenata | x | . | nb | . | sh | A | . | . | LH | s/l | ||
| Colydiidae | Colydium elongatum | x | 3 | nb | . | s | F | ! | . | L/N | s | ||
| Corylophidae | Arthrolips obscura | x | 2 | nb | . | s | A | . | . | LH | s | ||
| Corylophidae | Arthrolips nana | x | x | 2 | nb | . | ss | P | . | . | LH | s | |
| Corylophidae | Orthoperus atomus | x | . | nb | . | s | P | . | . | LH | s/l | ||
| Corylophidae | Orthoperus corticalis | x | . | nb | . | s | P | . | . | LH | l | ||
| Endomychidae | Symbiotes latus | x | 2 | nb | . | ss | M | ! | . | LH | s/l | ||
| Endomychidae | Symbiotes gibberosus | x | x | 2 | nb | . | s | M | ! | . | Bu | s | |
| Sphindidae | Sphindus dubius | x | . | nb | . | s | P | . | . | LH | s/l | ||
| Ciidae | Rhopalodontus perforatus | x | 3 | nb | . | h | P | . | . | LH | s/l | ||
| Ciidae | Sulcacis nitidus | x | x | . | nb | . | sh | P | . | . | Bu | s/l | |
| Ciidae | Sulcacis bidentulus | x | 2 | nb | . | ss | P | . | . | LH | s/l | ||
| Ciidae | Cis castaneus | x | . | nb | . | sh | P | . | . | L/N | s/l | ||
| Ciidae | Cis micans | x | x | x | . | nb | . | sh | P | . | . | LH | s/l |
| Ciidae | Cis boleti | x | . | nb | . | sh | P | . | . | L/N | s/l | ||
| Ciidae | Cis rugulosus | x | . | nb | . | mh | P | . | . | LH | s/l | ||
| Ciidae | Cis fagi | x | x | . | nb | . | h | P | . | . | LH | s/l | |
| Ciidae | Cis fusciclavis | x | x | . | nb | . | mh | P | . | . | LH | s/l | |
| Ciidae | Orthocis alni | x | . | nb | . | h | P | . | . | LH | s/l | ||
| Ciidae | Cis vestitus | x | 2 | nb | . | mh | P | . | . | LH | s/l | ||
| Ciidae | Ennearthron cornutum | x | . | nb | . | sh | P | . | . | LH | s/l | ||
| Bostrichidae | Lyctus cavicollis | x | . | nb | . | ss | A | . | . | ei | s | ||
| Bostrichidae | Xylopertha retusa | x | 3 | nb | . | ss | F | ! | . | Ei | l | ||
| Anobiidae | Ptinomorphus imperialis | x | x | . | nb | . | h | A | . | . | LH | s/l | |
| Anobiidae | Dryophilus pusillus | x | x | . | nb | . | mh | F | . | . | NH | s/l | |
| Anobiidae | Xestobium rufovillosum | x | x | x | . | nb | . | h | A | . | . | LH | s/l |
| Anobiidae | Hyperisus plumbeum | x | x | . | nb | . | h | A | . | . | LH | s | |
| Anobiidae | Ernobius mollis | x | . | nb | . | h | A | . | . | NH | s | ||
| Anobiidae | Oligomerus brunneus | x | x | x | 3 | nb | . | mh | A | . | . | LH | s/l |
| Anobiidae | Hemicoelus canaliculatus | x | . | nb | . | mh | A | . | . | LH | s | ||
| Anobiidae | Priobium carpini | x | . | nb | . | s | A | . | . | Ei | s | ||
| Anobiidae | Ptilinus pectinicornis | x | x | x | . | nb | . | sh | A | . | . | LH | s |
| Anobiidae | Ptilinus fuscus | x | . | nb | . | s | A | . | . | LH | s | ||
| Anobiidae | Xyletinus ater | x | . | nb | . | ss | A | . | . | Ei | s | ||
| Anobiidae | Xyletinus fibyensis | x | 2 | nb | . | ss | A | . | . | Ei | s | ||
| Anobiidae | Xyletinus vaederoeensis | x | x | 2 | nb | . | ss | A | . | . | Zit | s | |
| Anobiidae | Dorcatoma flavicornis | x | 3 | nb | . | s | A | ! | . | LH | s | ||
| Anobiidae | Dorcatoma chrysomelina | x | x | x | 3 | nb | . | h | A | ! | . | Ei | s |
| Anobiidae | Dorcatoma substriata | x | 2 | nb | . | ss | P | ! | . | LH | s | ||
| Anobiidae | Dorcatoma minor | x | x | nb | nb | . | ss | P | . | . | LH | s | |
| Anobiidae | Dorcatoma androgyna | x | nb | nb | . | E17 | P | . | . | LH | s | ||
| Anobiidae | Dorcatoma dresdensis | x | 3 | nb | . | h | P | . | . | LH | s | ||
| Anobiidae | Dorcatoma robusta | x | x | 2 | nb | . | mh | P | ! | . | LH | s | |
| Ptinidae | Ptinus rufipes | x | . | nb | . | h | A | . | . | LH | s/l | ||
| Ptinidae | Ptinus sexpunctatus | x | 3 | nb | . | s | xS | . | . | LH | s | ||
| Oedemeridae | Ischnomera cyanea | x | . | nb | . | s | A | . | . | LH | s/l | ||
| Salpingidae | Lissodema cursor | x | . | nb | . | mh | F | . | . | LH | l | ||
| Salpingidae | Rabocerus gabrieli | x | 2 | nb | . | s | F | ! | . | LH | s | ||
| Salpingidae | Salpingus planirostris | x | x | . | nb | . | sh | F | . | . | L/N | s/l | |
| Salpingidae | Salpingus ruficollis | x | x | . | nb | . | h | F | . | . | LH | s/l | |
| Pyrochroidae | Pyrochroa coccinea | x | . | nb | . | sh | A | . | . | LH | s/l | ||
| Pyrochroidae | Schizotus pectinicornis | x | x | . | nb | . | h | A | . | . | LH | s/l | |
| Scraptiidae | Scraptia fuscula | x | x | x | 3 | nb | . | mh | A | . | . | Ei | s |
| Scraptiidae | Anaspis frontalis | x | x | x | . | nb | . | sh | A | . | . | LH | s/l |
| Scraptiidae | Anaspis maculata | x | . | nb | . | ss | A | . | . | LH | s/l | ||
| Scraptiidae | Anaspis thoracica | x | . | nb | . | sh | A | . | . | LH | s/l | ||
| Scraptiidae | Anaspis rufilabris | x | x | . | nb | . | sh | A | . | . | LH | s/l | |
| Scraptiidae | Anaspis flava | x | x | x | . | nb | . | sh | A | . | . | LH | s |
| Aderidae | Aderus populneus | x | x | 3 | nb | . | s | M | . | . | Ei | s | |
| Aderidae | Euglenes oculatus | x | 2 | nb | . | s | M | ! | . | L/N | s | ||
| Mordellidae | Tomoxia bucephala | x | x | x | . | nb | . | h | A | . | . | Bu | s |
| Mordellidae | Mordella brachyura | x | . | nb | . | sh | A | . | . | LH | s/l | ||
| Mordellidae | Mordellistena neuwaldeggiana | x | . | nb | . | h | A | . | . | LH | s/l | ||
| Mordellidae | Mordellistena variegata | x | . | nb | . | h | A | . | . | LH | s/l | ||
| Melandryidae | Orchesia micans | x | . | nb | . | mh | P | . | . | LH | l | ||
| Melandryidae | Orchesia fasciata | x | 3 | nb | . | ss | P | . | . | LN | l | ||
| Melandryidae | Orchesia undulata | x | x | . | nb | . | mh | P | . | . | LH | s/l | |
| Melandryidae | Anisoxya fuscula | x | 3 | nb | . | s | P | . | . | LH | s/l | ||
| Melandryidae | Serropalpus barbatus | x | . | nb | . | s | A | . | . | NH | s/l | ||
| Melandryidae | Melandrya caraboides | x | 3 | nb | . | s | A | . | . | LH | s/l | ||
| Melandryidae | Conopalpus testaceus | x | x | x | . | nb | . | mh | A | . | . | LH | l |
| Tetratomidae | Tetratoma fungorum | x | x | . | nb | . | mh | P | . | . | LH | s | |
| Alleculidae | Allecula morio | x | x | x | 3 | nb | . | mh | M | . | . | Ei | s |
| Alleculidae | Prionychus ater | x | x | 3 | nb | . | mh | M | . | . | LH | s | |
| Alleculidae | Pseudocistela ceramboides | x | x | 2 | nb | . | s | M | . | . | Ei | s/l | |
| Alleculidae | Mycetochara maura | x | x | x | . | nb | . | h | A | . | . | LH | s |
| Tenebrionidae | Bolitophagus reticulatus | x | x | 3 | nb | . | h | P | . | . | Bu | s/l | |
| Tenebrionidae | Eledona agricola | x | . | nb | . | mh | P | . | . | LH | s/l | ||
| Tenebrionidae | Diaperis boleti | x | . | nb | . | h | P | . | . | LH | s/l | ||
| Tenebrionidae | Platydema violaceum | x | 3 | nb | . | s | P | . | . | LH | s | ||
| Tenebrionidae | Pentaphyllus testaceus | x | 3 | nb | . | mh | A | . | . | LH | s | ||
| Tenebrionidae | Corticeus unicolor | x | x | . | nb | . | sh | A | . | . | LH | s/l | |
| Tenebrionidae | Corticeus bicolor | x | x | x | 3 | nb | . | mh | F | . | . | L/N | s |
| Tenebrionidae | Corticeus fasciatus | x | 2 | nb | . | s | A | ! | 2 | Ei | s | ||
| Tenebrionidae | Uloma culinaris | x | x | x | 2 | nb | . | mh | A | ! | . | L/N | s/l |
| Tenebrionidae | Neatus picipes | x | 1 | nb | . | mh | A | ! | 1 | LH | s | ||
| Tenebrionidae | Stenomax aeneus | x | x | . | nb | . | h | A | . | . | L/N | s/l | |
| Scarabaeidae | Cetonia aurata aurata | x | x | . | . | § | h | A | . | . | LH | s/l | |
| Scarabaeidae | Protaetia fieberi | x | 2 | 1 | § | ss | M | ! | . | Apf | s | ||
| Scarabaeidae | Protaetia marmorata | x | x | 2 | 3 | § | mh | M | ! | . | Bu | s | |
| Scarabaeidae | Osmoderma eremita | x | x | 2 | 2 | § | mh | M | ! | 2 | LH | s | |
| Scarabaeidae | Gnorimus nobilis | x | x | x | 3 | 2 | § | s | M | ! | . | LH | s/l |
| Lucanidae | Platycerus caraboides | x | x | . | 3 | § | s | A | . | . | LH | s | |
| Lucanidae | Sinodendron cylindricum | x | x | 3 | 4 | § | h | A | . | . | LH | s/l | |
| Cerambycidae | Spondylis buprestoides | x | . | . | § | mh | A | . | . | NH | s/l | ||
| Cerambycidae | Arhopalus rusticus | x | . | . | § | mh | A | . | . | NH | s | ||
| Cerambycidae | Grammoptera ustulata | x | x | . | 2 | § | ss | A | . | . | LH | s/l | |
| Cerambycidae | Grammoptera ruficornis | x | . | . | § | sh | A | . | . | LH | s/l | ||
| Cerambycidae | Grammoptera abdominalis | x | . | 4 | § | s | A | . | . | Ei | s | ||
| Cerambycidae | Rutpela maculata | x | . | . | § | sh | A | . | . | LH | s/l | ||
| Cerambycidae | Pedostrangalia revestita | x | 2 | 3 | § | ss | A | . | . | LH | s/l | ||
| Cerambycidae | Cerambyx scopolii | x | 3 | 2 | § | s | F | . | . | LH | s/l | ||
| Cerambycidae | Axinopalpis gracilis | x | 1 | 1 | § | s | F | . | . | Ei | s/l | ||
| Cerambycidae | Obrium cantharinum | x | x | 2 | 1 | § | ss | F | . | . | LH | s | |
| Cerambycidae | Ropalopus femoratus | x | 3 | 1 | § | ss | F | . | . | LH | s | ||
| Cerambycidae | Phymatodes testaceus | x | . | . | § | h | F | . | . | LH | l | ||
| Cerambycidae | Poecilium alni | x | . | 2 | § | s | F | . | . | Ei | s/l | ||
| Cerambycidae | Xylotrechus rusticus | x | 2 | 1 | § | s | F | . | . | LH | s/l | ||
| Cerambycidae | Xylotrechus antilope | x | . | 1 | § | s | F | . | . | LH | s/l | ||
| Cerambycidae | Plagionotus detritus | x | 2 | 3 | § | s | F | ! | . | Ei | s | ||
| Cerambycidae | Plagionotus arcuatus | x | . | . | § | s | F | . | . | LH | s/l | ||
| Cerambycidae | Anaglyptus mysticus | x | . | . | § | mh | F | . | . | LH | s/l | ||
| Cerambycidae | Leiopus nebulosus | x | x | . | . | § | sh | F | . | . | LH | s/l | |
| Cerambycidae | Exocentrus adspersus | x | 3 | 1 | § | s | F | . | . | Ei | s | ||
| Cerambycidae | Exocentrus lusitanus | x | 3 | 3 | § | s | F | . | . | Li | s/l | ||
| Cerambycidae | Saperda perforata | x | 2 | 2 | § | ss | F | ! | . | Pa | s | ||
| Cerambycidae | Tetrops praeustus | x | . | . | § | h | F | . | . | LH | s/l | ||
| Anthribidae | Tropideres albirostris | x | x | 3 | nb | . | s | A | . | . | LH | s | |
| Anthribidae | Pseudeuparius sepicola | x | . | nb | . | s | A | . | . | LH | s/l | ||
| Anthribidae | Platystomos albinus | x | . | nb | . | mh | A | . | . | LH | s/l | ||
| Curculionidae | Scolytus rugulosus | x | x | . | nb | . | mh | F | . | . | LH | s | |
| Curculionidae | Scolytus intricatus | x | x | . | nb | . | sh | F | . | . | LH | s/l | |
| Curculionidae | Scolytus mali | x | . | nb | . | mh | F | . | . | LH | s | ||
| Curculionidae | Scolytus carpini | x | . | nb | . | mh | F | . | . | LH | l | ||
| Curculionidae | Scolytus multistriatus | x | x | . | nb | . | h | F | . | . | Ul | s | |
| Curculionidae | Phloeotribus spinulosus | x | . | nb | . | mh | F | . | . | NH | l | ||
| Curculionidae | Hylastes ater | x | . | nb | . | mh | F | . | . | Ki | l | ||
| Curculionidae | Hylastes opacus | x | . | nb | . | mh | F | . | . | Ki | s/l | ||
| Curculionidae | Hylurgops palliatus | x | . | nb | . | h | F | . | . | NH | l | ||
| Curculionidae | Polygraphus grandiclava | x | . | nb | . | s | F | . | . | LH | s | ||
| Curculionidae | Hylesinus crenatus | x | . | nb | . | s | F | . | . | Es | s | ||
| Curculionidae | Hylesinus toranio | x | . | nb | . | s | F | . | . | Es | s | ||
| Curculionidae | Hylesinus varius | x | x | x | . | nb | . | sh | F | . | . | Es | s/l |
| Curculionidae | Dryocoetes villosus villosus | x | . | nb | . | mh | F | . | . | Ei | s | ||
| Curculionidae | Trypophloeus binodulus | x | x | 3 | nb | . | ss | F | . | . | Pap | s | |
| Curculionidae | Ernoporicus fagi | x | x | . | nb | . | h | F | . | . | LH | s/l | |
| Curculionidae | Ernoporus tiliae | x | x | . | nb | . | h | F | . | . | LH | s/l | |
| Curculionidae | Pityophthorus pubescens | x | . | nb | . | s | F | . | . | Ki | s | ||
| Curculionidae | Pityophthorus lichtensteini | x | . | nb | . | s | F | . | . | NH | s/l | ||
| Curculionidae | Pityophthorus glabratus | x | . | nb | . | mh | F | . | . | Ki | s | ||
| Curculionidae | Taphrorychus bicolor | x | x | x | . | nb | . | h | F | . | . | LH | s/l |
| Curculionidae | Pityogenes chalcographus | x | . | nb | . | sh | F | . | . | NH | s/l | ||
| Curculionidae | Xyleborinus saxesenii | x | x | x | . | nb | . | sh | F | . | . | L/N | s/l |
| Curculionidae | Xyleborus monographus | x | x | x | . | nb | . | h | F | . | . | LH | s |
| Curculionidae | Xyleborus dryographus | x | . | nb | . | ss | F | . | . | LH | s/l | ||
| Curculionidae | Cyclorhipidion bodoanus | x | . | nb | . | ss | F | . | . | LH | s/l | ||
| Curculionidae | Trypodendron domesticus | x | x | . | nb | . | h | F | . | . | LH | s/l | |
| Curculionidae | Trypodendron lineatus | x | . | nb | . | h | F | . | . | LH | s/l | ||
| Curculionidae | Platypus cylindrus | x | 3 | nb | . | ss | F | . | . | LH | s/l | ||
| Curculionidae | Rhyncolus punctatulus | x | x | 2 | nb | . | s | A | . | . | Roß | s | |
| Curculionidae | Phloeophagus lignarius | x | . | nb | . | s | A | . | . | LH | s/l | ||
| Curculionidae | Stereocorynes truncorum | x | x | x | . | nb | . | h | A | . | . | LH | s/l |
| Curculionidae | Magdalis ruficornis | x | . | nb | . | h | F | . | . | LH | s | ||
| Curculionidae | Magdalis flavicornis | x | . | nb | . | mh | F | . | . | Ei | s | ||
| Curculionidae | Magdalis cerasi | x | . | nb | . | h | F | . | . | LH | s | ||
| Curculionidae | Magdalis armigera | x | . | nb | . | mh | F | . | . | Ul | s | ||
| Curculionidae | Magdalis caucasica | x | 1 | nb | . | ss | F | . | . | Ul | s | ||
| Curculionidae | Magdalis phlegmatica | x | . | nb | . | mh | F | . | . | Ki | s | ||
| Curculionidae | Hylobius abietis | x | . | nb | . | h | F | . | . | NH | s/l | ||
| Curculionidae | Echinodera hypocrita | x | . | nb | . | mh | A | . | . | LH | s/l |
RLDt: Rote Listen von Deutschland (GEISER 1998) Gefährdungskategorien: 0 = Ausgestorben/Ausgerottet/Verschollen; 1 = vom Aussterben bedroht; 2 = stark gefährdet; 3 = gefährdet; . = derzeit keine Gefährdung erkennbar; RLSn: Rote Listen von Sachsen (GEBERT 2009: Laufkäfer; KLAUSNITZER 1994: Bockkäfer und KLAUSNITZER 1995: Blatthornkäfer und Hirschkäfer): nb = nicht bearbeitet; . = derzeit keine Gefährdung erkennbar) BV: Bundesartenschutzverordnung H = Häufigkeit: ss = sehr selten; s = selten; mh = mäßig häufig; h = häufig; sh = sehr häufig; (subjektive Einschätzung des Autors bezogen nur auf Sachsen und auf 30jährige empirische Erfahrungen auf Grundlage vieler Gutachten und Untersuchungen sowie einer 100.000 Datensätze umfassenden Datenbank mit mehr als 3.200 Käferarten von mehr als 1.700 Fundorten in Sachsen); ÖG = Ökologische Gilde (nach SCHMIDL & BUSSLER 2003) = Substratgilden: F = Frischholzbesiedler, A = Altholzbesieder, P = an Pilzen, die auf Holz wachsen und M = Mulmhöhlenbesiedler; xS = Arten mit xylobionten Sonderbiologien, z.B. Baumsaftfresser, Kommensalen in Nestern anderer Holzinsekten) s = sonstige Arten mit Relevanz für morsches Lindenholz; IA = Indikatorarten bzw. waldökologisch besonders relevante Arten (v.a. unter Berücksichtigung historischer Aspekte wie Faunentradition, die auf Bestandeskontinuität fußt) (nach SCHMIDL & BUSSLER 2003); UR = Urwald-Reliktart nach MÜLLER et al. (2005): Kategorie 1 (Urwaldrelikte im engeren Sinne) und Kategorie 2 (Urwaldrelikt-Arten im weiteren Sinne) mit zusätzlichen Anforderungen an Requisiten, Ressourcen und Strukturen, wie z.B. große Waldflächen, seltene Holzpilze, starke Totholzdimensionen, hohes Baumalter, Heliophilie der Bestände, lange Verweildauer bzw. späte Sukzessionsstadien der Holzstruktur im Abbauprozess); BA= Baumart: Apf = Apfel; Bu = Buche; Ei = Eiche; Es = Esche; Fi = Fichte; Hbu = Hainbuche; Ki = Kiefer; LH = Laubholz; L/N = Laub- und Nadelholz; NH = Nadelholz; Li = Linde; Pa = Pappel; Roß = Rosskastanie; Ul = Ulme; Zi = Zitterpappel; BE= Exposition der Totholzstruktur: s = an Stehend-Totholz; l = an Liegend-Totholz; s/l = keine Bevorzugung der Exposition;